Photobiology is the study of the interactions of light (technically non-ionizing only) with living organisms. The most widespread interaction is photosynthesis, the process which converts light into the chemical energy that drives the majority of life on earth. We'll be limiting our discussion to plants but photobiology occurs ubiquitously throughout life forms.

Light is a form of energy that propagates via electromagnetic waves. These waves have different wavelengths and frequencies which affect the amount of energy carried by each light packet and types of particles the packets can interact with. Our eyes are sensitive to light that has wavelengths between around 400-800nm (1 nanometer, abbreviated nm, equals 1 x 10⁻⁹ meters). Coincidentally, the process of photosynthesis also relies on light in a similar range of wavelengths.
Packets of light are called photons and can be thought of as particles without mass. The energy of each photon depends on the wavelength and frequency of the light with the variables related via the equation, Ephoton = hf = hc/λ, where h is plank's constant equal to 6.6261 x 10^(-34) Js, f is frequency in cycles per second (Hz), c is the speed of light equal to 3 x 10⁸ meters/second and λ is the wavelength of the lightwave specified in meters.

For visible wavelengths, photons are emitted when electrons fall from a higher energy level to a lower energy level. The initial energy that raises an electron into an excited state is supplied by an external source. This can be nuclear reactions (like in the sun!), electrical excitation, thermal excitation or collisions. Electrical voltage from a wall socket is the most common input energy for artificial lighting. The electrical voltage can drive direct excitation of electrons (like in LEDs), ionize gases (like in fluorescents) or it can heat up an element which then emits photons (like in incandescent lighting).
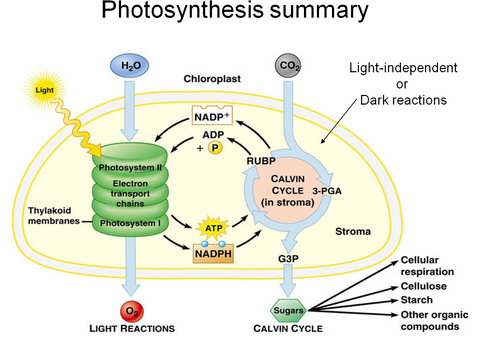
Photosynthesis occurs in plant cell organelles known as chloroplasts. It involves 2 separate but coupled reactions, known as the light reaction and the dark reaction (Calvin cycle). The light reaction requires light and water (emitting O₂) which drives a cascade of ultra-fast electron transfers that creates an electro-chemical gradient across the thylakoid membranes. This electro-chemical gradient drives chemo-mechanical reactions which phosphorylate ADP to ATP (a universal energy storage molecule) and reduce NADP+ to NADPH. These energy molecules are then transported to the stroma where they undergo the Calvin cycle and use their newly acquired energy states to transform carbon dioxide from the air into sugars. The sugars can then be used for cellular respiration or converted to polymers where they can be used for longer term energy storage or converted into structural elements.

Photon absorption in the light reaction takes place on antenna complexes comprised of an array of pigments. These pigments can include chlorophyll (a),(b), β-carotene, lycopene and others. Each pigment has its own absorption spectrum profile (see below). It should be noted that the antenna complexes have a funneling structure where photons with higher energies are absorbed by pigments on the outside of the complex and then the energy is transferred to molecules closer to the reaction center in a cascade. In the reaction center, the energy is converted into intermediary chemical states that are more stable than the short-lived excited states among pigment molecules.
Sometimes photo-damage can occur in the antenna complexes due to overexposure from light. There exists multiple "healing" pathways which scavenge free radicals that could potentially lead to damage. However, in conditions of overexposure, the scavenging pathways simply cannot keep up. Repair for damage in the areas is very costly from a molecular energetics perspective and thus can lead plants unequipped to deal with higher light intensities. The result is overexposed plants can suffer and or perish.
Above are the isolated absorption curves for different different pigments of the antenna complex. Of particular interest, are the two peaks of chlorophyll (a) and (b) in the blue and red. Many early LEDs grow lights were initially engineered to target these two peaks and this created a purple look. The logic here was that since plants are green, they reflect all the green light instead of using it for photosynthesis.

However, the action spectra of photosynthesis has a very different in shape than the individual pigments. Above is a graph of the relative rate at which photons yield photosynthetic activity (McCree, K.J. et al, 1972a). The curve was established by evaluating the input (CO₂) and byproduct (O₂) of photosynthesis in a controlled environment while plants were illuminated at given wavelengths. Note that this graph is the relative quantum yield per photon with the peak being around 625nm. This response differs from the individual pigment absorption of chlorophyll due to a number of physiological factors. Firstly, chlorophylls are not the only pigments responsible for photosynthesis and other pigments and cartenoids widen the range of absorption of the antenna complexes. Secondly, although green light may not be absorbed by antenna complex pigments as efficiency at the leaf surface, it is able to penetrate deeper into the photosynthetic tissue and is absorbed by chloroplasts on the backside of the leaf as well as leaves potentially below the initial leaf of incidence.
Photosynthesis is not the only way plants use light. Lighting spectra has an affect on plant growth habit. This is a highly species dependent trait. However, the general understanding is that blue light can inhibit cell expansion which leads to reduced leaf expansion in some plants and/or less stem etiolation. The result is often more compact plants. On the flip side, far red photons have the opposite effect, leading to enhanced cell expansion and increased leaf area and/or stem elongation depending on the species.
Although not tied to a specific photobiology effect, the importance of white light for plant growth cannot be overemphasized. An old Japanese proverb mentions "The Footsteps of the Farmer are His Best Fertilizer". White light provides the human eye the ability to diagnose pest, disease and nutrient issues.
Navigation List
References
Blankenship, Robert E. Molecular Mechanisms of Photosynthesis. Oxford: Blackwell Science, 2002. Print.
Blankenship RE, Tiede DM, Barber J, et al. Comparing photosynthetic and photovoltaic efficiencies and recognizing the potential for improvement. Science. 2011;332(6031):805-809.
Both, A. J. Measuring LED Lighting Systems and Developing Guidelines for Evaluation, Comparison and Use. Rep. SCRI‐LED, 11 June 2013. Web. 12 Feb. 2014.
Bugbee, Bruce. 2020. Apogee Instruments. https://www.apogeeinstruments.com/videos-and-tutorials. (01-01-2020)
Ellison AM, Farnsworth EJ. 2008. Prey availability directly affects physiology, growth, nutrient allocation and scaling relationships among leaf traits in 10 carnivorous plant species. Journal of Ecology 96: 213–221.
Green, M. A., Emery, K., Hishikawa, Y., Warta, W. and Dunlop, E. D. (2014), Solar cell efficiency tables (version 43). Prog. Photovolt: Res. Appl., 22: 1–9.
Khan, M. Nisa. Understanding LED Illumination. Boca Raton: CRC, 2014. Print.
Koning, Ross E. 1994. Light. Plant Physiology Information Website. http://plantphys.info/plant_physiology/light.shtml. (8-13-2014)
Koshel, R. John. Illumination Engineering: Design with Nonimaging Optics. Piscataway, NJ: IEEE, 2013. Print.
Kyte, Lydiane, John Kleyn, Holly Scroggins, and Mark Bridgen. Plants From Test Tubes. Portland:Timber Press, 2013. Print.
McCree, K.J. (1972a) Action Spectrum, Absorptance and Quantum Yield of Photosynthesis in Crop Plants. Agricultural Meteorology, 9, 191-216.
McCree, K.J. (1972b) Test of Current Definitions of Photosynthetically Active Radiation against Leaf Photosynthesis Data. Agricultural Meteorology, 10, 443-453.
Mitchell, Cary A., A. J. Both, C. M. Bourget, John F. Burr, Chieri Kubota, Roberto G. Lopez, Robert C. Morrow, and Erik S. Runkle. "LEDs: The Future of Greenhouse Lighting!" Chronica HORTICULTURAE 52.1 (2012): 6-12. Print.
Mitchell, Cary A. Developing LED Lighting Technologies and Practices for Sustainable Specialty-Crop Production. Rep. NIFA SCRI, 15 July 2012. Web. 12 Feb. 2014.
Narukawa, Yukio, et al. “White light emitting diodes with super-high luminous efficacy.” J. Phys. D: Appl. Phys. 43 (2010) 354002 (6pp).
Ross, J. and M. Sulev. 2000. Sources of errors in measurements of PAR. Agricultural and Forest Meteorology 100, 103-125.
Shenzhen Runlite Technology Co., Ltd., “Runlite Epistar,” SMD 5050 Series Data Sheet, Feb. 2014
Singhal, G. S., G. Renger, S.K. Sopory, K.D. Irrgang, and Govindjee. Concepts in Photobiology: Photosynthesis and Photomorphogenesis. Boston: Kluwer Academic, 1999. Print.
Torres, Ariana P., Christopher J. Currey, and Roberto G. Lopez. "Getting The Most Out Of Light Measurements." Greenhouse Grower (2010): 46-54. Issue. 27 Aug. 2010. Web. 14 Feb. 2014.
Wu, Nancy. "High Brightness Led Tube Light Fixtures Bulbs Replacement T8 8ft 2400mm 35W." - Quality LED Tube Light Fixtures for Sale. Shenzhen Greelife Technology Co., Ltd., 2012. Web. 15 July 2014.
Žukauskas, Artūras, Michael Shur, and Remis Gaska. Introduction to Solid-state Lighting. New York: J. Wiley, 2002. Print.
